The resting membrane potential
of large nerve fibers when not transmitting nerve signals is about –90
millivolts. That is, the potential inside the fiber is 90 millivolts more
negative than the potential in the extracellular fluid on the outside of the fiber.
In the next few paragraphs, we explain all the factors that determine the level
of this resting potential, but before doing so, we must describe the transport
properties of the resting nerve membrane for sodium and potassium.
Active Transport of Sodium and
Potassium Ions through the Membrane—the Sodium-Potassium (Na+ -K+) Pump. First,
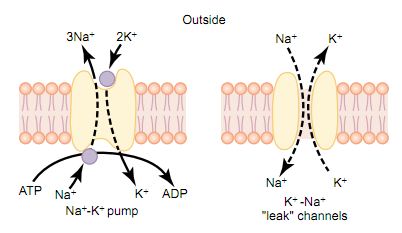
let us recall that all cell membranes of the body have a powerful Na+ -K+ that
continually pumps sodium ions to the outside of the cell and potassium ions to
the inside, as illustrated on the left-hand side in. Further, note that this is
an electrogenic pump because more positive charges are pumped to the outside
than to the inside (three Na+ ions to the outside for each two K+ ions to the
inside), leaving a net deficit of positive ions on the inside; this causes a
negative potential inside the cell membrane The Na+ -K+ also causes large
concentration gradients for sodium and potassium across the resting nerve
membrane Leakage of Potassium and Sodium Through the Nerve Membrane. The right
side of Figure below shows a channel protein in the nerve membrane through which
potassium and sodium ions can leak, called a potassium-sodium (K+ -Na+) “leak”
channel. The emphasis is on potassium leakage because, on average, the channels
are far more permeable to potassium than to sodium, normally about 100 times as
permeable. As discussed later, this differential in permeability is exceedingly
important in determining the level of the normal resting membrane potential.
Origin of the Normal Resting Membrane Potential
The figure shows the important
factors in the establishment of the normal resting membrane potential of–90
millivolts. They are as follows.
1.
Contribution of the
Potassium Diffusion Potential.
In the figure below1A, we make
the assumption that the only movement of ions through the membrane is diffusion
of potassium ions, as demonstrated by the open channels between the potassium
symbols (K+) inside and outside the membrane. Because of the high ratio of
potassium ions inside to outside, 35:1, the Nernst potential corresponding to
this ratio is –94 millivolts because the logarithm of 35 is 1.54, and these
times –61millivolts is –94 millivolts. Therefore, if potassium ions were the
only factor causing the resting potential, the resting potential inside the fiber
would be equal to –94 millivolts, as shown in the figure.
2.
Contribution of Sodium
Diffusion through the Nerve Membrane.
The figure shows the addition of slight
permeability of the nerve membrane to sodium ions, caused by the minute
diffusion of sodium ions through the K+ -Na leak channels. The ratio of sodium
ions from inside to outside the membrane is 0.1, and this gives a calculated Nernst
potential for the inside of the membrane of +61 millivolts. But also shown in
Figure 1B is the Nernst potential for potassium diffusion of –94 millivolts. How
do these interact with each other, and what will be the summated potential?
This can be answered by using the Goldman equation described previously. Intuitively,
one can see that if the membrane is highly permeable to potassium but only
slightly permeable to sodium, it is logical that the diffusion of potassium con
tributes far more to the membrane potential than does the diffusion of sodium.
In the normal nerve fiber, the permeability of the membrane to potassium is
about 100 times as great as its permeability to sodium. Using this value in the
Goldman equation gives a potential inside the membrane of –86 millivolts, which
is near the potassium potential shown in the figure.

3.
Contribution of the Na
-K Pump.
In Figure 1C, the Na+ -K+ pump
is shown to provide an additional contribution to the resting potential. In
this figure, there is continuous pumping of three sodium ions to the outside for
each two potassium ions pumped to the inside of the membrane. The fact that
more sodium ions are being pumped to the outside than potassium to the inside
causes continual loss of positive charges from inside the membrane; this
creates an additional degree of negativity (about –4 millivolts additional) on the
inside beyond that which can be accounted for by diffusion alone. Therefore, as
shown in Figure 1C, the net membrane potential with all these factors operative
at the same time is about –90 millivolts.
In summary, the diffusion
potentials alone caused by potassium and sodium diffusion would give a membrane
potential of about –86 millivolts, almost all of this being determined by
potassium diffusion. Then, an additional
–4 millivolts is contributed to the membrane potential by the
continuously acting electrogenic Na+ -K+ pump, giving a net membrane potential of
–90 millivolts.


